This weekend we were delighted to present our research at the annual Swansea Science Festival. It was a great opportunity to talk about microbes, sponges and microbial communities to kids, parents and attendees of all ages. We even had some visitors from the Star Wars world who wanted to know more about microbiomes!
Our stand was full of engaging activities from soft toys that looked like microbes and kids could sort into handcrafted sponges to build-your-own microbiome crafts for children!


We also had microscopes with an assortment of slides that everybody could use to have a look at the microscopic structures of plant, animals, and even bacteria, our invisible friends that do so much for our survival and the wellbeing of our planet.
For those more interested in the science behind it all we also had informative flyers to take home detailing how we go from thinking about microbes and marine sponges to abstracting those associations into concepts of evolution, metacommunities and symbioses. We then built up all the way to a 3rd tier of complexity linking those concepts to mathematical equations and models that help us reason about these complex, yet fundamental ecosystems that persist all around us and without which life on Earth would not be possible.
At the end of the day we got a wall full of handcrafted microbiomes from all the kids that visited our exhibit. All in all it was great fun!

The Leverhulme Trust supported our stand at the Science Festival through Research Project Grant RPG-2022-114 to ML.
Thanks to Gui, Jooyoung and Olivia for all your help. And thanks to the organisers, and Delyth in particular, for a great festival.
In early September, the first UK Mathematical Biology Conference took place at the University of Birmingham. Our lab was represented by Gui Araujo, who presented a poster showcasing our microbiome assembly framework.
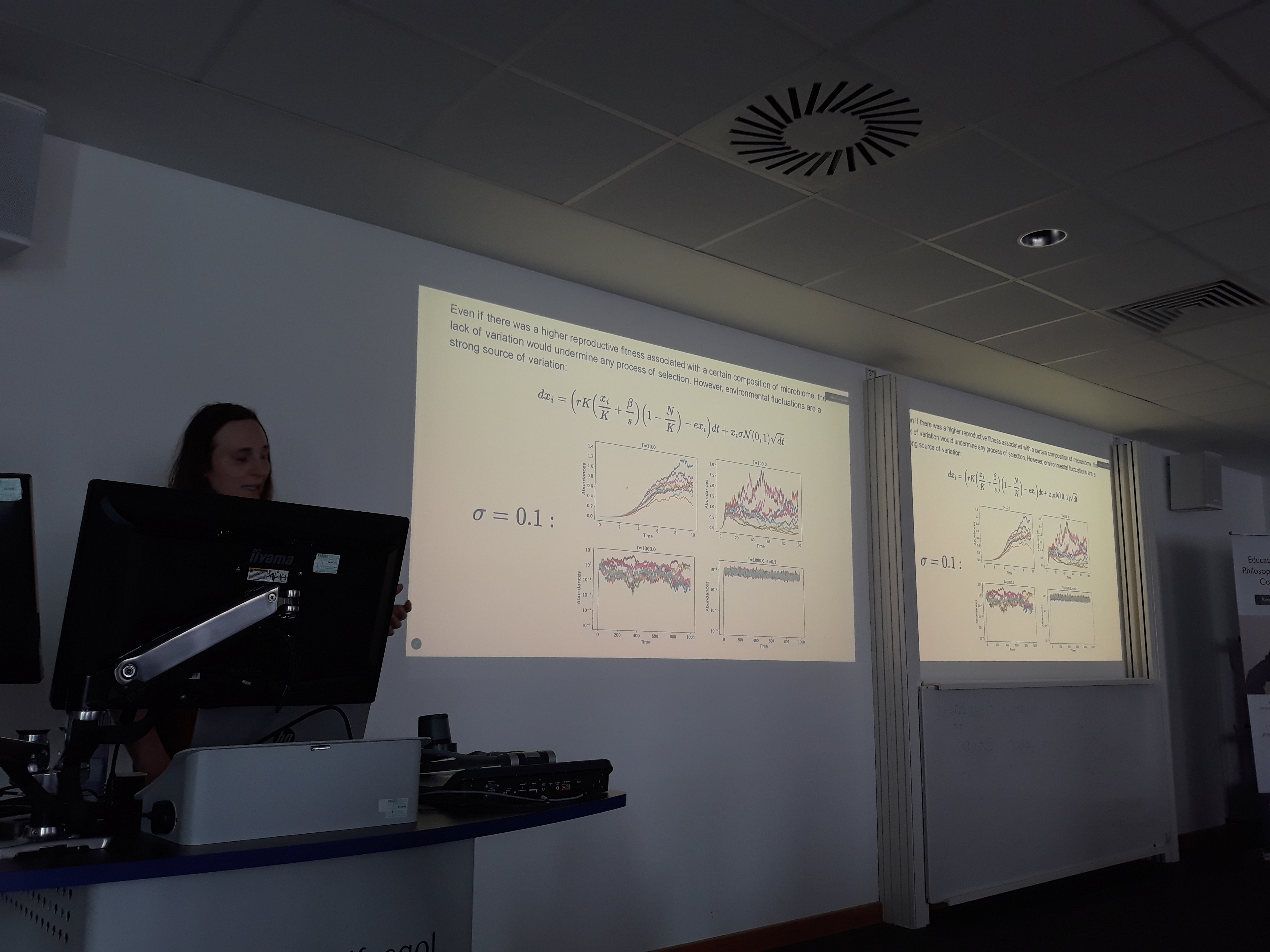
The presentation was structured in two parts. The first focused on powerful methods from stochastic calculus that substantiate the connection between 1) specific events creating and destroying individuals and 2) global changes in densities represented by continuous flows. Many researchers at the conference work with these tools, which provided a natural entry point to discuss our extensions of the population dynamics framework. The individual-population connection is particularly relevant for the mechanistic modelling of ecological populations, since equations such as the classic Lotka-Volterra can only be directly linked to dynamics at the level of real individuals through a connection like this one. It also serves as the first step of the connections explored in our microbiome assembly framework , ultimately going through the multilayer route of individual -> population -> community -> metacommunity.
The idea behind the connection is to first establish the individual-level dynamics as a Markov jump process, described through a master equation, by modelling knowledge about events as state-dependent chances of occurrence per time step. Then, an expansion is performed on the size of the system, which increases the scope without changing the structure, going from a discrete probabilistic description to a continuous deterministic one.
The second part of the presentation featured our framework and the model we developed within it to describe microbiome assembly. This model highlights the interplay of three mechanisms: microbial dispersal, enrichment within the host environment, and microbe–resource interactions. We used this model to describe richness and abundance patterns observed in marine sponge data. Sponge species are classified into two broad categories relating to their microbiome, sponges with high and low microbial abundance (HMA and LMA). HMA and LMA sponge species differ in all three mechanisms as a consequence of their differences in pumping rates, host selection, and degree of resource allocation from host to microbes. In our (upcoming) work, we compared the model simulations with real sponge data and showed a substantial agreement between the two.
This conference was a great opportunity to engage with other mathematicians/physicists working in several fields of biology in the UK and think about how interesting it is that the same methods can be used to investigate so many different aspects of life sciences.
Participation in this conference was made possible thanks to the support of the Leverhulme Trust.

In this tutorial, Gui goes through the theoretical reasoning and calculations involved in the process of parameter estimation of a standard linear regression, starting from Bayes equation.
Here, we will derive the process of linear regression, starting from a dataset and then calculating point estimations of the parameters of a linear model using the data. Suppose we obtain a series of $N$ data points containing joint measurements of two variables, $(x_i,y_i)$, with $i=1,2,…,N$. Then, we offer a theory, a model, of how these measurements should relate to each other. In our theory, we assume that the following is true:
\[y = ax+b.\]
For this particular experiment, we don’t know the specific values of $a$ and $b$. In statistical terminology, our theory has selected a model, but has not determined it. The estimation of $a$ and $b$ that are best suited to the data we obtained is called a model determination process, as is any process of parameter estimation. The relation between $y$ and $x$ could be described by another equation, but our theory currently assumes that this is the right relation.
In order to connect data with theory, we must go through Bayes equation, which states:
\[P(Theory|Data) = \frac{P(Data|Theory)P(Theory)}{P(Data)}.\]
The logical inversion stating that
\[P(Theory|Data) \propto P(Data|Theory)\]
is a deep and powerful statement that arguably substantiates science as a whole, because it allows the conversion from knowledge about empirical observations into knowledge about our theories. However, it comes with a strong caveat: scientific theories are (of course) subjected to empirical observations, and the very usage of empirical observations is intrinsically subjected to the existence of theories. In other words, you have to give to receive. Data means nothing unless it is positioned within a theoretical frame (which, by the way, is intrinsically subjective).
So far, our theory says that $y = ax+b$ is the relation between $y$ and $x$. However, when it comes to the actual data points $y_i$ and $x_i$, they do not follow exactly this relation. The problem is: we do not get to know the true values of these variables. There are many sources of errors and limitations in our observations of these variables and, even if our model is true, we do not have access to exact measurements. Therefore, we assume that the relation in the data differs from the expected model by an error. Thus, an updated model has to be applied to the data (and not to abstract $x$ and $y$ variables), and it involves the existence of the error:
\[y_i = ax_i+b+e_i,\]
where $e_i$ is the error. In the same way as we characterised the relation between the variables (as linear) by selecting a model, we also have to characterise the error by selecting a model for its distribution. This is also a part of the assumed theory, and it can be inferred from the dataset. In the standard linear regression, on top of the linear model, we assume a normal distribution of errors (which means that this is a procedure suited for a dataset following both the facts that the variables are linearly related and the observation errors are normally distributed). Thus, we have:
\[p(e) = N(e|0,\sigma^2) = \frac{1}{\sqrt{2\pi\sigma^2}}exp\Big({-\frac{e^2}{2\sigma^2}}\Big).\]
Now, with a fully determined model, we can instantiate the Bayes equation and the statistical process:
\[P\Big(a,b,\sigma\Big|\{x_i,y_i\}\Big) = \frac{P\Big(\{x_i,y_i\}\Big|a,b,\sigma\Big)P\Big(a,b,\sigma\Big)}{P\Big(\{x_i,y_i\}\Big)}.\]
In some way, we have to compute an estimation of the parameters $[a,b,\sigma]$. However, it is necessary to bypass the dreadful factor $P\Big({x_i,y_i}\Big)$. If we follow the Bayesian route, we boldly decide to use the full posterior. The way to bypass the probability of data (or normalisation factor) is by costly sampling the posterior using tricks. The standard is an MCMC algorithm, which samples from the posterior using only kernel ratios (thus cancelling out the normalisation factor).
The classic frequentist route for the linear regression is to bypass the normalisation through an optimisation process. Instead of obtaining the full posterior, we assume uninformative (unstructured) priors and determine the mode of the posterior. This gives us the point estimation of the parameters with maximum probability given the data. In this situation, the mode of the posterior is also the mode of the likelihood, which means maximising the probability of data given the model. The likelihood is easy to calculate and easy to maximise, therefore the resulting task is to just obtain the maximum likelihood. So we start by calculating the likelihood and forgetting everything else.
The first step is to assume that the set of observed pairs ${x_i,y_i}$ is composed of independent and identically distributed points. Therefore:
\[P\Big(\{x_i,y_i\}\Big|a,b,\sigma\Big) = \prod_{i=1}^NP\Big(x_i,y_i\Big|a,b,\sigma\Big)\]
Assuming our model $y_i = ax_i+b+e_i$, we have an expression for $y_i$ given the model and also given the measurement $x_i$. Then, we can decompose each factor using the definition of conditionals:
\[P\Big(x_i,y_i\Big|a,b,\sigma\Big) = P\Big(y_i\Big|x_i,a,b,\sigma\Big)P(x_i).\]
Then, all factors $P(x_i)$ can be left out as well, since they do not depend on the model parameters (and, if we also decompose the normalisation factor in the same way, these probabilities are cancelled out). Now, the probability of $y_i$ given the other variables is given directly by the model. If the error was zero, then this probability would be $1$ if $y_i=ax_i+b$ and zero otherwise. Since we have a normally distributed error, this probability is then normally distributed around the deterministic part of the model (by definition, the error is the distribution around the deterministic value, which is then a mean):
\[P\Big(y_i\Big|x_i,a,b,\sigma\Big) = \mathcal{N}(y_i|ax_i+b,\sigma^2) = \frac{1}{\sqrt{2\pi\sigma^2}}exp\Big({-\frac{(y_i-ax_i-b)^2}{2\sigma^2}}\Big).\]
Now, substituting back into the full likelihood, we have:
\[P\Big(\{x_i,y_i\}\Big|a,b,\sigma\Big)\propto\prod_{i=1}^N \frac{1}{\sqrt{2\pi\sigma^2}}exp\Big({-\frac{(y_i-ax_i-b)^2}{2\sigma^2}}\Big)=L.\]
The structure of this function is almost begging us to work with a log transformation. Luckily, the function $log(f)$ is monotonic on $f$, so the maximum of $f$ is also a maximum of $log(f)$. Therefore, if we maximise $log(L)$, the result will be the same, and it is a much easier task. Then:
\[log(L) = \sum_{i=1}^N \Big( -log(\sqrt{2\pi \sigma^2}) -\frac{(y_i-ax_i-b)^2}{2\sigma^2} \Big)\]
\[log(L) = -Nlog(\sqrt{2\pi}) - Nlog(\sigma) - \sum_{i=1}^N \frac{(y_i-ax_i-b)^2}{2\sigma^2}\]
This is the final expression for the log-likelihood. Now, we perform the maximisation in relation to all parameters by deriving in relation to each of them and equating to zero. All expressions must be valid at the same time (consider all derivatives below as partial derivatives):
First, in relation to $a$:
\[\frac{dlog(L)}{da} = 0 = \frac{d}{da} \Big( \sum_{i=1}^N (y_i-ax_i-b)^2 \Big)\]
Then, in relation to $b$:
\[\frac{dlog(L)}{db} = 0 = \frac{d}{db} \Big( \sum_{i=1}^N (y_i-ax_i-b)^2 \Big)\]
These two equations are actually independent of omega, so they can be solved independently as a 2D system. Then, the obtained values of $a$ and $b$ can be used to determine $\sigma$ in:
\[\frac{dlog(L)}{d\sigma} = 0 = -\frac{N}{\sigma} + \frac{1}{\sigma^3}\sum_{i=1}^N (y_i-ax_i-b)^2,\]
which translates into simply
\[\sigma^2 = \frac{1}{N}\sum_{i=1}^N (y_i-ax_i-b)^2.\]
By solving the equations above for $a$, $b$, and $\sigma$, we arrive at the following expressions. First for $a$ explicitly in terms of data points:
\[a = \frac{N \sum_i (x_i y_i) - \sum_i x_i \sum_i y_i}{N \sum_i x_i^2 - (\sum_i x_i)^2}\]
Then $b$:
\[b = \frac{ \sum_i y_i \sum_i x_i^2 - \sum_i x_i \sum_i (x_i y_i) }{ N \sum_i x_i^2 - (\sum_i x_i)^2 }\]
These expressions can be prettier if we summarise them in terms of the averages of the $y_i$’s and $x_i$’s. Then, we can use $a$ and $b$ to calculate $\sigma$ from its previous equation. However, there’s an additional detail. Since we are estimating 2 parameters in this case, that expression becomes a biased estimator of $\sigma$. For an unbiased estimation, we must discount the 2 degrees of freedom from the variance, and the expression then becomes:
\[\sigma = \sqrt{ \frac{1}{N - 2} \sum_{i=1}^N (y_i - ax_i-b)^2 }\]
If we run a standard linear regression as a statistical test, for example the function lm() in R, we are not only calculating the point estimates of the parameters as above. The parameter $\sigma$ is reported as a residual standard error (RSE), and R also includes estimations of the errors on the parameters that are calculated from the estimated $\sigma$, the parameter standard errors. We are also performing t-tests for the null hypotheses of $a$ or $b$ being equal to zero, and that is where p-values enter. Diagnostic statistics of residuals (such as their distribution summaries) are also provided to assess model assumptions like normality and homoscedasticity.